Histology of the eye and eyelid
A. Eyelid
Slide UCSF 173 eyelid H&E [DigitalScope] (virtual slide courtesy of the University of California, San Francisco)
- conjunctiva
- accessory glands (focus particularly on their secretory products):
- apocrine glands (of Moll)
- tarsal glands (of Meibohm)
- lacrimal glands (of Wolfring and of Krause, don't worry so much about which lacrimal gland is which, but you should be able to identify lacrimal glands in general and know what they secrete)
- muscles (focus on function and innervation):
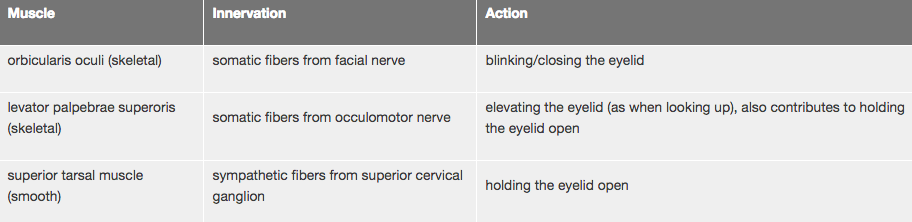
- orbicularis oculi
- levator palpebrae superioris (mostly only the tendon is visible although some strands of actual muscle can be seen)
- superior tarsal muscle of Müller
The eyelid or palpebra functions primarily to protect the the eye from mechanical damage and to help adjust the amount of incoming light. In addition, blinking the eyelid helps spread a liquid film of tears over the cornea (a structure discussed below) which, being avascular, is critically dependent upon the tear film as a means of gas exchange. Tears also contain lysozyme and other antibacterial proteins that help protect the cornea and conjunctiva from infection. The outer surface of the eyelid is covered by thin skin (epidermis) with eyelashes and their associated sebaceous glands (glands of Zeis) concentrated toward the margin of the lid. Also in this area are modified, coiled apocrine sweat glands (glands of Moll) that add secretory IgA, lysozyme, and other antibacterial agents to the tear film. At the margin of the lid is the muco-cutaneous junction of the outer epidermis and the conjunctiva of the eyelid which is a stratified columnar epithelium containing goblet cells that contribute mucous secretions to the tear film. The conjunctiva continues along the interior surface of the eyelid and then makes a sharp turn at a region known as the "fornix" to reflect onto the surface of the eye where it then becomes continuous with the outer epithelium of the cornea.
The two major components forming the core of the eyelid are the orbicularis oculi muscle and the tarsal plate. The orbicularis oculi muscleis a muscle of facial expression (innervated by the facial nerve) consisting of circularly-oriented skeletal muscle fibers overlying the tarsal plate. The tarsal plate consists of dense fibroelastic tissue in which additional "accessory" glands are found that add secretions to the tear film. The most significant of these glands are the tarsal glands (of Meibohm), which are linear arrays of sebaceous glands connected to central ducts that open at the muco-cutaneous junction of the eyelid. There are about 20 or such glands in each of the upper and lower eyelids and their sebaceous secretion (along with sebaceous glands associated with the eyelashes) contributes a critical lipid component to the tear film that reduces surface tension and retards evaporation. In the upper part of the tarsal plate are some small accessory lacrimal glands (of Wolfring) that, like the main lacrimal gland (which we won't study, but you will see in your dissection of the orbital region in the anatomy lab), are serous glands that contribute much of the watery and proteinaceous components of the tear film including antibacterial agents such as lysozyme. Additional accessory lacrimal glands (of Krause) can be observed at the fornix.
Here is a summary of the glands of the eyelid and their contributions to the tear film:

Attached to the tarsal plate are two muscles that serve to elevate the eyelid. One of these muscles is the levator palpebrae superioris which is a skeletal muscle (innervated by the occulomotor nerve). This muscle is not readily visible in your section, but you can see its dense connective tissue tendon running between the orbicularis oculi muscle and the tarsal plate. If you follow the tendon to the top of the section, you should be able to see some of the skeletal muscle fibers. The other muscle is the superior tarsal muscle (of Müller) which is smooth muscle (innervated by postganglionic sympathetic fibers from the superior cervical ganglion) that can be seen in this section just above the tendon of the levator palpebrae at the fornix. If you trace the course of the smooth muscle fibers, you'll see that they follow the curvature of the conjunctiva and insert into the upper margin of the tarsal plate (near the lacrimal glands of Wolfring). As these two muscles are the only means by which the eyelids are held open, lesions affecting either the occulomotor nerve or sympathetic innervation (e.g. Horner's syndrome) will result in a noticeable drooping of the eyelid, a condition known as ptosis.
Here is a summary of the muscles of the eyelid, their innervation, and function:
B. Corneal/Scleral Layer of the eye (Grants Figs. 7.37, 7.38; Netter Figs. 83, 84, 86A, 86B)
Slide EYE-2 eye H&E [DigitalScope]
The sclera is the opaque part of the outermost coat or tunic of the eye (the tunica externa) that covers about 5/6 of the eye. The anteriormost 1/6 of the tunica externa is optically transparent and is known as the cornea, which is comprised of three cellular layers and two noncellular layers. The outermost layer is the corneal epithelium which is a stratified, non-keratinized epithelium that is continuous with the conjunctival epithelium overlying the adjacent sclera. Deep to the epithelium is Bowman’s membrane which is the basement membrane of the corneal epithelium. Underneath Bowman’s membrane is the connective tissue of the corneal stroma which makes up much of the cornea. The stroma contains regular, orthogonally arranged bundles of type I collagen critical to the optical properties of the cornea. Deep to the stroma is Descemet’s membrane which is the basement membrane of the corneal endothelium. The corneal endothelium is a simple squamous epithelium facing the anterior chamber of the eye. Transparency of the cornea requires precise control of the hydration of the stroma and it is cells of the corneal endothelium that perform this function. Unlike the corneal epithelium, corneal endothelial cells have very limited proliferative potential, so severe damage to this epithelium can only be repaired by transplantation.
The limbus is the region where the cornea transitions to sclera and you’ll see this is also where the cornea meets with the tissue of the iris at what is known as the irideo-corneal angle. Within the connective tissue of the angle lies a network of endothelial lined channels which is the trabecular meshwork, and deep to the meshwork within the connective tissue of the sclera is the venous canal of Schlemm. Fluid of the aqueous humor produced by the ciliary body (discussed below) in the posterior chamber is absorbed within this meshwork and is drained via the canal of Schlemm into episcleral veins thus maintaining intraocular pressure at around 15 mm Hg. What structural problems in the eye can lead to glaucoma?
C. Chambers and Lens of the eye (G 7.37, 7.38; N 83, 84, 86A, 86B)
There are three recognized chambers of the eye:
- anterior chamber, the space between the cornea and the iris
- posterior chamber, the space between the posterior surface of the iris and the anterior surface of the lens.
- vitreous chamber (or cavity), the space between the posterior surface of lens and the retina
The anterior and posterior chambers contain aqueous humor, which is a watery fluid produced by the ciliary body (discussed below) whereas the vitreous chamber contains the gelatinous vitreous body which is comprised of collagen type II fibers suspended within a highly hydrated gel of hyaluronan, extracellular matrix, and water.
Separating the aqueous chambers from the vitreous chamber is the lens. The lens (particularly the core) is very difficult to section so it is frequently damaged during preparation of microscope slides. However, there is are several features that can still be seen. On the anterior surface (the side facing the direction of incoming light) is the lens capsule, which is actually the basement membrane of the underlying lens epithelium which consists of a layer of epithelial cells that are more squamous toward to the anterior apex of the lens and become taller as you move posteriorly toward the lens equator. At the lens equator, the lens epithlelial cells differentiate into lens cells that elongate and rotate such that they extend from the anterior aspect of the lens to the posterior surface of the lens. As new cells are added peripherally, older cells are pushed inward. The cells eventually become entirely filled with lens crystallin proteins and lose their nuclei as they are pushed toward the core of the lens, at which point they are called lens fibers. What are cataracts and how do they form?
Upon observation of the areas peripheral to the equator of the lens, you should note the presence of delicate collagenous strands. The fibers constitute what is collectively known as the suspensory ligament or zonule (of Zinn) which runs from the equator of the lens to the ciliary body (discussed below). These fibers anchor the lens in space and allow the ciliary body to affect the curvature of the lens.
D. Uveal/Choroidal Layer of the eye (G 7.37, 7.38; N 83, 84, 86A, 86B)
1. lris
- Iridial stroma with its “uncovered” anterior surface and melanocytes [DigitalScope]
- Sphincter (constrictor) pupillae muscle [DigitalScope]
- Pigmented epithelium and cytoplasm of the dilator pupillae muscle [DigitalScope]
The iris controls the amount of light that enters through the pupil and divides the anterior chamber from the posterior chamber. On the side facing the anterior chamber is the connective tissue of the irideal stroma. Note that it is not covered by any sort of epithelium and that it consists of a loose connective tissue containing fibroblasts and a variable number of melanocytes. At the pupillary margin of the iris is a band of circularly arranged smooth muscle (so cut in cross section here) known as the sphincter or constrictor pupillae muscle. This muscle receives postganglionic parasympathetic innervation from the ciliary ganglion (the preganglionic fibers originate from the accessory occulomotor nucleus (of Edinger-Westphal), and its contraction causes the pupil to constrict. Continuing around the pupillary margin, you should note that the portion of the iris facing the posterior chamber consists of two prominent cell layers, the most obvious being the posterior pigmented epithelium. The pigment serves the obvious function of blocking light such that only that coming through the pupil is focused through the lens onto the retina. Just above the pigmented epithelium is a slightly less pigmented anterior epithelium which actually consists of myoepithelial cells that are specialized such that they project their contractile portions (the eosinophilic layer just above the pigmented portion) in a radial manner so that they cause the pupil to dilate when they contract. Thus, this muscular portion of the anterior epithelium constitutes the dilator pupillae muscle, and you should recall that this muscle receives postganglionic sympathetic innervation from the superior cervical ganglion. Note that these two muscles are in a constant “tug of war” such that inhibition of sympathetic input (e.g. Horner’s syndrome) will result in obvious constriction of the pupils (a condition known as “miosis”) whereas inhibition of parasympathetic input (such as treatment with eye drops containing a parasympathetic antagonist like atropine) will result in dilation of the pupils (a.k.a. “myadriasis”).
2. Ciliary Body
The ciliary body functions primarily to control the shape of the lens and produce aqueous humor. Just behind the iris, the surface of the ciliary body is thrown into folds known as ciliary processes. The innermost cells facing the “lumen” of the eyeball are non-pigmented and are ultimately continuous with the ganglion cell layer of the neural retina, however these cells are obviously NOT neural. Instead, you should note that they are cuboidal and, particularly along the ciliary processes, quite eosinophilic which is due to their high content of mitochondria and ion channels essential for their primary role in the production of aqueous humor. Recall that aqueous humor produced here flows from the posterior chamber through the opening at the iris into the anterior chamber and is then drained via the trabecular meshwork and the canal of Schlemm. Deep to the non-pigmented epithelium is a layer of pigmented cells which is ultimately continuous with the pigmented cells of the visual (or “neural”) retina. Above the pigmented epithelium is a rich vascular bed which is ultimately continuous with the choriocapillary layer of the neural retina. This vascular supply is particularly necessary for metabolic support of the aqueous humor-producing cells and the ciliary muscle, discussed below.
The bulk of the ciliary body consists of longitudinally, radially, and circularly arranged bundles of smooth muscle which is collectively known as the ciliary muscle within what is also technically part of the choroid layer. Like the sphincter pupillae, the ciliary muscle also receives postganglionic parasympathetic innervation from the ciliary ganglion (the preganglionic fibers originate from the accessory occulomotor nucleus (of Edinger-Westphal). During the process known as accommodation, the circularly oriented muscle fibers contract, thus constricting this ring of muscle. This, in turn, releases tension on the suspensory ligaments (or zonules of Zinn) that run from the ends of the ciliary processes to the equator of the lens thus causing the lens to become more spherical to increase its focal power necessary for near vision. Because the sphincter pupillae is also innervated by the same parasympatheitc fibers, the pupil will also constrict during this process thus increasing the “depth of field” which also helps for near vision.
- Melanocyte layer
- Choriocapillary layer
- Glassy (Bruch’s) membrane (a thickened “basement membrane” composed of the basal laminae of the pigment epithelial cells and choroid capillary endothelium plus some intervening choroid elastic and collagen fibrils)
The choroid is an element of the tunica vasculosa and consists of three obvious layers. Closest to the connective tissue sclera is a layer of pigmented melanocytes. Next is an extensive capillary bed of the choriocapillary layer followed by Bruch’s membrane, which is a common basement membrane shared by the capillary endothelial cells and the adjacent pigmented epithelium of the retina (discussed below). The cells of the “outer retina” (e.g. the rod and cone photoreceptors) receive metabolic support from the choriocapillaris via exchange of materials across this basement membrane.
E. Retina (G 7.37, 7.38; N 83, 84, 86A, 86B)
This line marks the termination of the photoreceptive cells of the retina anteriorly. Anterior to this point, the “retina” appears just as a two layered structure with an inner, non-pigmented columnar-cuboidal epithelium and an outer layer of pigmented epithelium that forms the lining of the ciliary body and posterior surface of the iris. (Think of the embryology. The pupillary margin of the iris marks the folded anterior edge of the developing optic cup. This may help you figure out where basement membranes ought to be.)
- retinal pigment epithelium (RPE)
- layer of rod and cone cells outer segments
- outer limiting membrane
- outer nuclear layer
- outer plexiform layer
- inner nuclear layer
- inner plexiform layer
- ganglion cell layer
- nerve fiber layer
- internal limiting membrane
The retina actually consists of two components: an outermost layer of retinal pigment epithelium (RPE), which is composed of single layer of cuboidal melanin-containing cells and the neural retina which is a multilayered structure containing photoreceptors as well as neurons and glia. In life, these two components are fused into what we typically call the retina, and it is subdivided into 10 recognizable layers.
As mentioned above, the outermost layer (closest to the choriocapillaris) is the retinal pigment epithelium which is a single layer of pigmented cells that absorb light, thus preventing stray light from inappropriately reflecting back onto the rods and cones which have their outer segments in close contact with the apices of the pigmented epithelial cells. Tight junctions between the RPE cells also establish a blood-retina barrier to regulate the exchange of materials from the blood to the retina. The next layer is the layer of rod and cone outer segments, which, for the rod cells, are cylindrical whereas for the cone cells these outer segments are conical. Recall that rods are more sensitive to light and thus are the receptors primarily used in periods of low light intensity, but the resulting image is monochromatic. Cones, on the other hand, are sensitive to specific wavelengths of light allowing you to discern colors and more detailed visual information, but they require more intense lighting.
After the rod and cone outer segments is the outer limiting membrane, which is a dense line formed by the junctional complexes between the rod and cone cells and the supportive Muller glia. It separates the outer segments which are rich in photosensitive pigments from the rest of the retina which functions primarily to integrate and process the signals initiated by the rod and cone cells. Next is the outer nuclear layer consisting of the somata and nuclei of rod and cone cells. In general, the rod cell nuclei are typically small and spherical and located at all levels of the layer whereas the cone cell nuclei are larger and more ovoid and usually located just to the inside of the outer limiting membrane. From this layer, the rod and cone cells project their processes into the outer plexiform layer where they synapse with neuronal elements from the inner nuclear layer which is a highly complex layer containing the cell bodies of bipolar cells, horizontal cells, amacrine cells, and Muller glia cells (these cannot be told apart in routine H&E sections, but you should be aware that they are in this layer). The Muller glia, of course, are the primary support cell of the neural retina whereas the other cells of this layer are neurons that perform the initial processing of visual information and then relay that on to the retinal ganglion cells. The relaying of this information takes place in the inner plexiform layer which contains the axons of bipolar and amacrine cells and the dendritic trees of the retinal ganglion cells, the somata and nuclei of which reside in the retinal ganglion cell layer. The cells of this layer are indeed sensory neurons of variable size (correlated with slightly different functional characteristics) that receive input from the amacrine and bipolar cells and send that along to other components of the visual system via axons that travel in the nerve fiber layer containing non-myelinated axons and supportive astroglia. These axons coalesce at the optic disc and then exit the eye via the optic nerve. The final element of the neural retina is a thin internal limiting membrane consisting of the expanded terminal portions of Müller glia and a basement membrane that delimits the neural retina from the vitreal cavity.
3. Location of blood vessels
Note that there is a dual blood supply for the retina with the outer layers supplied from vessels in the choriocapillaris and the inner layers supplied by retinal vessels that branch from the central retinal artery.
4. Where retinal detachment occurs
The retina develops from two opposing epithelia of the optic cup that eventually fuse into a single structure with the apical domain of the outer epithelium (the pigment epithelium) ultimately coming into contact with the apical domain of the inner neuroepithelium (rod and cone outer segments of the neural retina). As a result, the weakest point of the retina is at this interface of the pigment epithelium and the rod and cone cell outer segments and it is along this plane where “retinal detachment” usually occurs. Because the outer layers of neural retina are so dependent upon the pigment epithelium and choriocapillaris for metabolic support, the result of such detachment can be irreversible ischemia and necrosis of the rod and cone cells, thus causing permanent blindness.
5. Emergence of optic nerve (optic disc) [DigitalScope]
The point at which all of the axons from the retinal ganglion cells converge and exit the eye via the optic nerve is the optic disc. You should note that there is a break in the retina at this point, so this area is incapable of detecting light and therefore produces a blind spot in your lateral visual field. Click here for a demonstration of blind spot caused by the optic disc.
6. Fovea
Slide EYE-1 eye H&E [DigitalScope]
This slide is from a different sample in which the fovea may be seen. The fovea is a small shallow depression in the central region of the retina located such that most of the incident light collected by the cornea and lens is focused onto this region. Most of the inner layers of the retina are markedly reduced or absent and what dominates is a layer of photoreceptors composed entirely of cone cells that are more slender and rodlike than they are elsewhere to accommodate their dense packing. Peripheral to the fovea, you should note that the inner nuclear layer and ganglion cell layer is much thicker compared to other parts of the retina. This is because unlike other areas of the retina where ganglion cells may receive input from many photoreceptors, for the fovea, there is close to a 1:1 ratio of ganglion cells to photoreceptors to allow for very fine discrimination of colors and details. However, in order to allow light to pass unimpeded to the photoreceptors, the all of the associated ganglion cells and cells of the inner nuclear layer are heaped up on the sides of the fovea. Retinal vessels are also absent in the region of the fovea for the same reason.
Histology of the Ear (G 7.78a, 7.78b)
I. Bony Labyrinth
Embedded within the petrous portion of the temporal bone is the bony labyrinth which consists of the central vestibule, three semicircular canals, which arise and end at the vestibule, and the cochlea. It contains a fluid called perilymph, which has a composition similar to extracellular fluids (high in Na+, low in K+).
- Between the vestibule and middle ear is a tissue-covered hole in the bone, the “oval window”. The footplate of the stapes [DigitalScope] is attached to the oval window on the middle ear side. Here, vibrations of the tympanic membrane are converted into fluid vibrations of perilymph via the bones of the middle ear.
- The cochlea [DigitalScope], located anteriomedially to the vestibule consists of a tube spiraling around a bony axis (the modiolus).
B. Vestibulocochlear Nerve - cochlear division and vestibular division [slide EAR-2, DigitalScope]
The vestibulocochlear nerve (CN VIII) enters the petrous temporal bone to innervate the inner ear. The innervation of the cochlea and vestibule is as follows:
- The cell bodies of the cochlear neurons occupy a canal (the spiral canal) that spirals up the modiolus. This spiral aggregate of nerve cells is called the spiral (or cochlear) ganglion [DigitalScope]. These are bipolar sensory neurons: their dendrites synapse with the hair cells in the organ of Corti (discussed below) and their axons come together in the cochlear nerve, which occupies a central canal in the modiolus and exits at the base of the cochlea.
- In the sensory regions of the vestibule, hair cells of the maculae and cristae (discussed below) synapse with dendrites of a corresponding aggregation of sensory neurons in the vestibular (or Scarpa’s) ganglion [DigitalScope], which then send their axons through the vestibular nerve. The processes of the vestibular nerve join with those of the cochlear nerve to form the vestibulocochlear nerve (cranial nerve VIII) (orientation).
II. Membranous labyrinth - The membranous labyrinth consists of the interconnected utricle, saccule, three semicircular ducts, the endolymphatic duct and sac, and the cochlear duct. It contains a fluid, endolymph, much like intracellular fluid in composition (high K+, low Na+).
A. Otolith Organs - Macula of the Utricle and Saccule [slide EAR-2, DigitalScope]
The utricle and the saccule contain the otolith organs that lie within the bony vestibule (orientation) and serve to detect linear acceleration. These otolith organs consist of sensory areas called maculae, and you should note that they are oriented somewhat perpendicular to each other to be able detect movement in different planes. Taking a closer look at a macula [DigitalScope], you should be able to identify the sensory epithelium comprised of hair cells and supporting cells. It is overlaid by a gelatinous material (otolithic membrane) in which are embedded calcite crystals (otoliths or otoconia). The otoconia are quite visible in some slides, hardly visible in others. During linear acceleration, inertial drag of these crystals causes deflection of the hair cells and subsequent signaling into the CNS. Beneath the sensory epithelium is a delicate connective tissue (filled with perilymph) and nerve fibers from the bipolar neurons of the vestibular (Scarpa’s) ganglion.
B. Semicircular Canals and Cristae Ampullari [slide EAR-4, DigitalScope]
The semicircular ducts have dilations (ampullae) near their origins from the utricle. Each ampulla has a ridge of tissue, or crista ampullaris, [DigitalScope] that projects into the lumen. The crista has a sensory epithelium similar to that of the macula; recall, however, that the cristae are specialized to detect angular (or rotational) acceleration. The hairs of the hair cells project into a gelatinous material called the cupula. The cupula projects into the lumen more than the otolithic membrane of the macula, and lacks otoconia. Here, inertial drag of the perilymph pushes on the cupula and causes deflection of the hair cells and subsequent signaling into the CNS.
What would happen if any otoconia or other debris were to become lodged somewhere in a semicircular canal or in a cupula? Answer
Note that the “membrane” of the membranous labyrinth is suspended from the bone by a delicate connective tissue and that in the Ear-1 and Ear-2 slides this tissue has torn such that the membrane is now squished up against the crista, so there’s not much of the cupula that can be seen. (orientation)
C. Cochlear Duct and Organ of Corti
[slide EAR-1, DigitalScope]
[slide 80a, DigitalScope]
The cochlear duct [DigitalScope] or scala media contains the organ of Corti, which detects sound (hearing). The cochlear duct is a triangular tube that is suspended in the middle of the spiraling bony labyrinth of the cochlea, thus subdividing this space into three spiraling sub-compartments, or “scalae”: the scala vestibuli, which opens on the vestibule; the scala media; and the scala tympani, which ends at the round window (the round window is not seen in any your slides). The scala vestibuli and scala tympani are both elements of the bony labyrinth and contain perilymph; the scala media or cochlear duct is an element of the membranous labyrinth and contains endolymph.
Elements of the cochlear duct that can be seen in cross section are: (orientation)
- The vestibular membrane (also called Reissner’s membrane) is the tissue separating the cochlear duct from the scala vestibuli.
- The stria vascularis [DigitalScope] is a stratified epithelium along the outer wall of the cochlear duct that is unique in that it is vascularized (most epithelia are avascular) by an extensive capillary network. Cells of the stria vascularis are responsible for the production and maintenance of endolymph.
- The [DigitalScope] extends from the tip of the osseous spiral lamina of the central modiolus to the outer wall of the cochlea and separates the cochlear duct from the scala tympani. The organ of Corti rests on the basilar membrane. Notice that the width of the basilar membrane changes such that it is shorter at the base and longer toward the apex of the cochlea. What is the significance of this change in length?
- The organ of Corti [DigitalScope] consists of two kinds of hair cells and various supporting cells in a complex arrangement.
The organ of Corti contains: (orientation)
- The outer hair cells surrounded by outer phalangeal cells. There are three rows of outer hair cells. The apices of these cells and their phalangeal cells are joined together to form the reticular membrane (also called reticular lamina or apical cuticular plate) that separates endolymph in the scala media from underlying corticolymph and perilymph of the scala tympani. Lateral to the outer hair cells and phalangeal cells are other support cells, but you don’t need to worry about knowing their specific types. Note that outer hair cells account for only ~5-10% of the sensory input into the auditory system. The primary function of outer hair cells is actually to contract when stimulated, thus “pulling” on the tectoral membrane thereby stimulating the inner hair cells (see animation).
- Outer and inner pillar cells outline a triangular shaped tunnel, called the inner tunnel, which is filled with perilymph-like fluid called corticolymph.
- The inner hair cells are in a single row close to the inner pillar cells (you may see more than one inner cell nucleus because of the thickness of the section). Note that the inner hair cells account for ~90-95% of the sensory input into the auditory system.
- The organ of Corti is overlaid by a gelatinous tectorial membrane (produced and maintained by the columnar cells found atop the spiral limbus just medial to the organ of Corti).
- Nerve fibers enter the organ of Corti through openings in a shelf of bone extending from the modiolus like the thread of a screw. The nerve fibers pass between supporting cells to synapse with the hair cells. Compare the innervation and function of the inner vs. outer hair cells.
A few notes about deafness: Disruption of any part of the process by which sound waves are transduced into input into the auditory portion of the CNS will result in “deafness.” Damage to the eardrum or ossicles results in so-called “conduction” deafness whereby sound waves are no longer transmitted into the inner ear. In this instance, a patient would NOT be able to hear a tuning fork held near the pinna, and the loss of hearing would extend across the entire range of frequencies. However, placing the stem of the fork on a bony part of the skull (e.g. the mastoid process) would then transmit vibrations directly to the inner ear (via the bone) where they could then be “heard.”
Loss of components within the cochlea results in sensorineural deafness which is more frequency-specific (i.e. the patient will not be able to hear specific pitches depending on the location of the damage in the cochlea). Loss of OUTER HAIR CELLS in a particular region of the cochlea would result in a “threshold shift” whereby sound of a particular frequency could still be detected (because the inner hair cells are still intact), but it would have to be LOUDER to make up for the fact that there are no outer hair cells to help stimulate the inner hair cells. This type of hearing loss can be compensated by a hearing aid.
Loss of INNER HAIR CELLS in a particular region of the cochlea would result in an almost complete inability to detect specific frequencies regardless of how loud they are. Loss of SPIRAL GANGLION CELLS would have a similar effect since these are the cells that actually project into the CNS. In both cases, the deafness could only be corrected with a cochlear implant.
|